

La respuesta inmune frente a antígenos extraños requiere una coordinación perfecta de todas las células que participan en las diferentes fases de esa respuesta. El objetivo de la respuesta es la destrucción rápida de los microorganismos pero debe garantizar la mínima repercusión sobre la totalidad de las células y tejidos del organismo. Los linfocitos T , fundamentalmente, llevan a cabo la regulación de este proceso. Se denominan células colaboradoras a las células encargadas de coordinar la respuesta inicial frente a los patógenos, y se denomina células reguladoras a las células que velan por el respeto de la integridad de lo propio y, una vez controlada la infección, desmontan la respuesta. Se conocen 3 tipos de células colaboradoras que coordinan respuestas frente a parásitos intracelulares: el TH1 (linfocito T helper), el helmintos (TH2) y las bacterias de crecimiento extracelular y hongos (TH17). La hiperfunción de las TH17 está asociada a enfermedades como la artritis reumatoide debido a la hipersecreción de la citocina con mayor efecto proinflamatorio: la interleucina-17.
La condición de célula colaboradora o reguladora es actualmente objeto de revisión. Las células TH1, TH2 y TH17, además de colaboradoras, tienen funciones supresoras de las otras respuestas, ya que son mutuamente antagónicas. Igualmente se ha descrito muy recientemente que las células T reguladoras también tienen un papel importante en la coordinación de los primeros pasos de la infección vírica de modo directo y también indirectamente, induciendo, a través de la secreción del factor de crecimiento transformante β, la diferenciación de las TH17.
The immune response foreign antigens require a perfect coordination of cells that participate in its different phases. The objective of the response is the rapid destruction of the microorganisms with a minimum repercussion on self-cells and tissues. The regulation of this process is carried out fundamentally by T lymphocytes. There are two main types of coordinator cells: helper cells, what organize the initial immune response, and regulatory cells, what avoid immune attack against self and once the infection is controlled, disassemble the response. There are three types of helper cells which coordinate answers to intracellular parasites (TH1), helmints (TH2) and extracellular bacteria and fungi (TH17). The hyperfunction of TH17 cells is associated with diseases as reumatoid arthritis, due to the hypersecretion of the proinflammatory citoquine IL17. The condition of helper or regulatory cell is the current object of review. TH1, TH2 and TH17 cells have helper and also regulatory functions.
In addition, regulatory T cells play an important role in the coordination of the first moments of the response to viral infection in a direct and indirect way, inducing differentiation of TH17 cells.
El sistema inmune, a través de la inmunidad innata y la inmunidad adaptativa, es el encargado de proteger frente a agresiones de cualquier tipo (generalmente microbianas) que puedan afectar las células o los tejidos. La respuesta defensiva más eficaz es la que lleva a cabo la inmunidad adaptativa, que permite la identificación y eliminación de los microorganismos y moléculas extrañas a causa de la actividad coordinada de cientos de células con funcionalidades distintas.
El proceso de la respuesta adaptativa consta de 5 fases:
- •
Reconocimiento del antígeno extraño
- •
Identificación, activación y expansión de células con moléculas que reconocen específicamente a ese antígeno, dando lugar a clones antigenoespecíficos
- •
Diferenciación de las células en reposo a células con fenotipo efector
- •
Desarrollo de la respuesta propiamente dicha: las células o sus productos inactivan y eliminan a los patógenos
- •
Inactivación de las células efectoras una vez resuelta la situación
La puesta en marcha del proceso requiere una coordinación perfecta de todas las células que participan en las diferentes fases, de modo que la destrucción de los microorganismos sea rápida pero que garantice la mínima repercusión de esa respuesta sobre la totalidad de las células y tejidos del organismo.
En la década de 1970, los linfocitos T fueron las primeras células encargadas de la coordinación de la respuesta que se describieron. Ya a finales de esa década se los clasificaba en 3 subgrupos según su capacidad funcional: los linfocitos T citotóxicos (Tc), con capacidad efectora; los linfocitos T helper (colaboradores) (TH), encargados de coordinar la acción efectora de los diferentes tipos celulares, y los linfocitos T supresores (Ts) encargados de desmontar la respuesta inmune una vez vencida la infección.
La sinapsis inmunitariaEl proceso de reconocimiento de antígenos, activación e inicio de la actividad efectora de los linfocitos T tiene lugar en los primeros pasos de la respuesta inmune y requiere el intercambio de información entre la célula que ha identificado y atrapado al patógeno y el linfocito T. Este proceso tiene lugar en una estructura multimolecular formada entre ambas células, que se denomina sinapsis inmunitaria.
La célula presentadora de los antígenos (CPA) aporta información al linfocito T a través de diversos tipos de señales. Las señales esenciales son 3: la primera señal es el reconocimiento del antígeno por parte del receptor de la célula T, cuando le es presentado en las moléculas del complejo principal de histocompatibilidad (CPH) de la CPA; la segunda señal, o señal de conformidad, la recibe el linfocito T a través del antígeno CD28, que interactúa con las moléculas de la célula presentadora B7-1 y B7-2 (CD80/CD86), y la tercera señal está mediada por citocinas y determina el tipo de respuesta que se efectuará y la intensidad de ésta1,2.
Los linfocitos T colaboradoresDurante la década de 1980 se profundizó en el conocimiento de la funcionalidad de los linfocitos T y se identificaron 2 tipos de respuestas colaboradoras: la TH1 (inmunidad celular o retardada) y la TH2 (inmunidad humoral). Las TH1 son altamente efectivas en la eliminación de patógenos intracelulares y las TH2 son de gran importancia en la eliminación de microorganismos extracelulares y parásitos3.
Esta división en 2 subtipos se basó en el panel de citocinas que éstos eran capaces de secretar una vez activados, y con las que modulaban a diversos tipos celulares. Se denominó TH1 a los linfocitos secretores de interferón γ (IFN-γ) e interleucina 2 (IL-2), y se denominó TH2 a los linfocitos que liberan IL-4 e IL-134.
La diferenciación en TH1 o TH2 a partir de los linfocitos quiescentes se determina en la sinapsis inmunitaria, en función de las citocinas que están presentes y que funcionan como terceras señales durante el proceso de activación. La IL-12 promueve la transformación en células TH1 y la IL-4 promueve la transformación en células TH25. Hay un tercer tipo de células colaboradoras, las células TH3, que deprimen la respuesta (por esa razón no se tratan aquí, sino en el apartado de células reguladoras).
Nuevas células colaboradoras: los linfocitos T helper 17Esta clasificación de las células colaboradoras ha sido revisada actualmente debido a descubrimientos recientes, como la identificación de la familia de citocinas de la IL-17 y el estudio de sus funciones efectoras. La existencia de un tercer grupo de células reguladoras se dedujo tras comprobar que las células T tratadas con péptidos microbianos en presencia de Borrelia burgdorferi (el agente de la enfermedad de Lyme) producían IL-17; pero a esta citocina no la elaboran las TH1 ni las TH2, lo que implica la existencia de un nuevo subgrupo de linfocitos T (CD4+) que secretan IL-17 y que coordinan la respuesta inmune de un modo diferente a las TH1 o a las TH26.
A estas células se las denomina TH17, son el tercer tipo de células colaboradoras reconocido por la comunidad científica (tabla 1) y desempeñan un papel fundamental en la respuesta contra bacterias de crecimiento extracelular y hongos. Asimismo, se ha descrito para ellas un efecto proinflamatorio que les permite hacer de puente entre la inmunidad innata y la inmunidad adaptativa7,8.
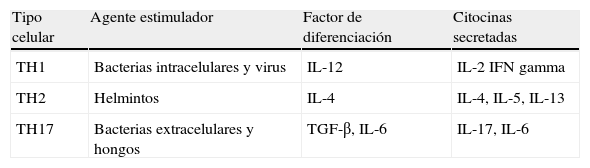
Células T colaboradoras
| Tipo celular | Agente estimulador | Factor de diferenciación | Citocinas secretadas |
| TH1 | Bacterias intracelulares y virus | IL-12 | IL-2 IFN gamma |
| TH2 | Helmintos | IL-4 | IL-4, IL-5, IL-13 |
| TH17 | Bacterias extracelulares y hongos | TGF-β, IL-6 | IL-17, IL-6 |
IFN: interferón; IL: interleucina; TGF-β: factor de crecimiento transformante β; TH: linfocito T helper.
Las citocinas implicadas en el control de la actividad TH17 son la IL-23, el factor de crecimiento transformante β (TGF-β) y la IL-6. El TGF-β y la IL-6 promueven la diferenciación de los linfocitos quiescentes en TH17 y, una vez diferenciados, la citocina que induce la proliferación de estas células es la IL-239.
Los diferentes tipos de células colaboradoras se inactivan mutuamente, de modo que los TH1 (a través del IFN-γ) inhiben selectivamente la actividad de los TH2 y los TH17. A su vez, los TH2 inhiben la proliferación de los TH1 y los TH17 mediante la IL-10 y la IL-4, y los TH17 inhiben a TH1 y a TH27,10,11.
La IL-17 es la primera de una familia de citocinas (familia de la IL-17). Actualmente se conocen 6 moléculas diferentes que se nombran desde IL-17A a IL-17F. La IL-17A es la más importante y por eso es a la que se denomina genéricamente como IL-17. El receptor para la IL-17A está presente en una amplia variedad de células y tejidos, tanto del sistema inmune (linfocitos B y T, monocitos, células de estirpe mieloide, estroma de médula ósea, etc.) como extrainmunes (epiteliales, fibroblastos, endotelio, etc.). Hay varios receptores similares al de la IL-17 aunque su función todavía no está bien definida. La deficiencia congénita del receptor de IL-17 en ratones conlleva que estos animales presenten una extrema sensibilidad a infecciones por gramnegativos y hongos12.
La IL-17A, al igual que la IL-17H, actúa sobre un amplio panel de células, y las estimula a secretar potentes mediadores de la inflamación como IL-1, TNF-α (tumor necrosis factor alpha ‘factor de necrosis tumoral alfa’), IL-6, IL-8, prostaglandina E2, quimiocinas y metaloproteasas. Además de actuar sobre las células del tejido, la IL-17 es esencial en el reclutamiento, activación y migración de otras células del sistema inmune13.
Células colaboradoras y enfermedadSe ha descrito que diversos procesos patológicos de base inmune se deben a disregulación de las células colaboradoras. Así, el exceso de las señales que generan los TH1 se asocia a procesos inflamatorios, mientras que el exceso de las señales que generan los TH2 desencadena enfermedad atópica, fundamentalmente alergias y asma14.
Aunque las células TH17 se han descubierto muy recientemente, su hiperfunción ya se ha asociado a procesos inflamatorios crónicos y autoinmunes promovidos, fundamentalmente, por el efecto proinflamatorio de la IL-1715. Se ha implicado a la IL-17 de modo directo en el desarrollo de diversas enfermedades autoinmunes, entre ellas destaca la artritis reumatoide (AR), enfermedad en la que se ha encontrado que la expresión de IL-17 está elevada en las zonas afectadas16.
En la AR, además de potenciar la actividad de IL-1 y TNF-α, la IL-17 tiene una acción directa en la evolución del cuadro clínico, puesto que estimula la diferenciación de los osteoclastos y promueve la destrucción de cartílago y hueso17,18.
Consecuentemente con estas observaciones, en modelos de AR en rata se ha descrito que la neutralización de la actividad de la IL-17, mediante el tratamiento con antagonistas de su receptor, atenúa la evolución de la enfermedad19.
Hasta hace unos años se consideraba que los principales causantes del daño tisular en las enfermedades autoinmunes eran las células TH1, pero actualmente se considera que las TH17 son las principales inductoras de enfermedad autoinmune: migran más rápidamente que las TH1 a las zonas de la lesión y, una vez allí, organizan la respuesta inflamatoria y son capaces de reclutar a otras células complementarias, entre ellas, las propias TH1, que necesariamente deben colaborar con las TH17 para que se produzca la inflamación y destrucción tisular15.
Otras células productoras de interleucina 17 competidoras con los linfocitos T helper 17Además de los TH17, se ha descrito que otros linfocitos T, como los gamma delta, son importantes secretores de IL-1720. Esto puede explicar el hecho paradójico de que en las articulaciones de los sujetos con AR donde las concentraciones de IL-17 son elevadas, las células predominantes son las TH1, que deberían inhibir la acción de la TH1721.
Células reguladorasEn experimentos realizados en la década de 1980 con diversos tipos de hibridomas, se identificó a un subtipo de células T que tenía la capacidad de suprimir las respuestas inmunes frente a antígenos específicos. Se observó esta capacidad supresora in vitro o in vivo y se la podía trasladar a cualquier individuo simplemente transfiriendo estas células, que se denominaron supresoras22.
No obstante, a finales de la década de 1980 y ante la falta de marcadores moleculares que permitieran identificarlas y estudiar su funcionamiento, las células supresoras fueron cayendo en el ostracismo y se llegó a descartar su existencia, pese a que nunca se pudo refutar el hecho en sí de la supresión. Las pocas referencias que algún inmunólogo hacía de estas células se etiquetaban de «paraciencia» e incluso se ridiculizaban23.
Ante el hecho evidente e irrefutable de la existencia de la tolerancia transferible, algunos científicos continuaron estudiándola, pero ya nadie se atrevía a usar la palabra «supresión», hasta el punto de llegar a referirse a ella sólo con la letra inicial para no nombrarla («Saying the ‘S’ word in public»)24. Estos estudios tuvieron su recompensa casi en la clandestinidad, en trabajos como el del grupo de Sakaguchi, que a mediados de la década de 1990 demostraron de modo definitivo la existencia de un subgrupo de células T especializadas en mantener la tolerancia a lo propio. En un modelo murino de autoinmunidad inducida tras timectomía, observaron que los ratones enfermos carecían de una población de linfocitos T CD4+CD25+ y se podía evitar la enfermedad si se les transfería este subgrupo de linfocitos T procedentes del bazo de otros animales que no eran autoinmunes (no presentaban timectomía)25,26.
Pese a que actualmente ya nadie duda de su papel esencial en el mantenimiento de la tolerancia a lo propio y en la inducción de tolerancia a los trasplantes, todavía se tiene el mismo problema que en la década de 1980: encontrar marcadores que identifiquen de modo inequívoco a las diversas poblaciones celulares con capacidad reguladora27.
Se ha descrito una amplia variedad de poblaciones de células reguladoras que controlan la respuesta inmune en diferentes puntos y a través de diversos mecanismos, su diversidad se ha llegado a comparar, incluso, con la variedad de sabores y olores28. Entre las células que poseen funciones reguladoras se encuentran el T CD4+, el T CD8+, el T gamma delta, los linfocitos citolíticos naturales (NK), los linfocitos T citolíticos naturales (NKT), el CD3+CD4−CD8− y los linfocitos B. Entre este «maremagno» de regulación, las células que más destacan (por ser las más estudiadas) son los linfocitos T CD4+ que expresan constitutivamente altas concentraciones de CD25+ y se las denomina genéricamente como células T reguladoras (Treg) (CD4+CD25high). Es conveniente recordar que la mayoría de los linfocitos T CD4+ quiescentes son constitutivamente CD25 negativos y sólo expresan este marcador (en concentración muy baja y de forma transitoria) en cuanto se activan tras el reconocimiento del antígeno (CD4+CD25low)29.
Es evidente que sólo con estos marcadores no es posible definir una población celular, por eso se han buscado otras moléculas adicionales que permitan identificar mejor a las células Treg. Se han descrito diversos marcadores presentes en las células Treg, como CD45RB, CD152, GITR, CD134 y CD62L, pero también se expresan en otros tipos celulares, por lo que son de poca utilidad. No obstante, el factor de transcripción FOXP3 sí parece ser muy característico de las células Treg30,31. Los sujetos que presentan mutaciones en el gen FOXP3 tienen el síndrome inmunodisregulación, poliendocrinopatía y enteropatía ligada al X (IPEX), una enfermedad que se caracteriza por presentar afectación intestinal y endocrina de origen autoinmune. Asimismo, los ratones con esta alteración genética (ratones scurfy) presentan un síndrome autoinmune letal. Tanto los enfermos como los ratones scurfy presentan concentraciones muy bajas de Treg en la sangre y las escasas células que poseen tienen una funcionalidad muy deficiente32. La función específica de FOXP3 no se conoce, aunque más bien parece ser indirecta a través de amplificar y estabilizar la transcripción de diversos genes específicos de regulación.
Aunque en ratones el modelo está muy claro, muy recientemente se ha puesto en duda que FOXP3 en humanos sea definitorio de las Treg, ya que también se encuentra presente en células de memoria. Como las células con mayor capacidad supresora que son FOXP3 positivas presentan en su membrana concentraciones muy bajas del receptor de IL-7 (CD127), se considera que el fenotipo CD4+ CD25+ CD127− es el mejor marcador de células Treg en humanos33.
Las células CD4+ reguladoras naturales e induciblesLa mayoría de las células Treg que se encuentran en sangre se originan en el timo y se las conoce como células Treg naturales. También, aunque hay otras células T CD4+ reguladoras inducibles que se originan en tejidos periféricos, las que mejor se conocen son las células T reguladoras inducibles (Tr1) y las TH3.
- •
Las Tr1 se diferencian a partir de CD4+CD25− tras la presentación del antígeno por células dendríticas inmaduras en presencia de citocinas como la IL-1034. Se localizan en tejidos inflamados y realizan su función supresora de una forma no dependiente del antígeno, a través de citocinas supresoras como IL-10 y TGF-â 35, mientras que las Treg se sitúan en los órganos linfoides periféricos y parecen actuar de forma antigenoespecífica a través de señales dependientes del contacto celular y de la secreción de IL35–37.
- •
Las células TH3 son células productoras de TGF-β con un funcionamiento muy similar al de las Tr138. Se las ha descrito en las placas de Peyer y los ganglios mesentéricos y se las considera como las causantes de los mecanismos de tolerancia oral39.
Como se ha visto con anterioridad, aunque las células más conocidas sean los linfocitos T CD4+ reguladores, hay varios tipos celulares con capacidad reguladora (los más conocidos se recogen en la tabla 2). Su existencia se ha deducido por datos funcionales, pero al igual que en el caso de las TR1 o TH3 no se conocen marcadores específicos que permitan distinguirlas de otras poblaciones celulares con capacidad colaboradora o citotóxica. Las principales células reguladoras se recogen en la tabla 2. Se citará por su importancia a:
- •
Los linfocitos T CD8+ supresores (CD8 reguladores, en palabras más «políticamente correctas»), que han vuelto a merecer atención por parte de la comunidad científica en los últimos años debido a un mejor conocimiento de su papel en procesos fisiológicos, como el mantenimiento de la tolerancia órganos inmunoprivilegiados, la tolerancia oral, y en situaciones especiales como procesos autoinmunes, trasplante de órganos y control de la enfermedad del injerto contra el huésped40,41. Las células T CD8+ tolerogénicas que mejor se conocen tienen el fenotipo CD8+CD28− y realizan su función a través del reconocimiento de antígenos en el CPH de las células dendríticas a las que convierten en tolerógenas mediante el bloqueo de la expresión de moléculas coestimuladoras en esas células dendríticas. Las CPA toleradas no son capaces de activar células T CD4+ helper42.
- •
Las células NKT que se definen por ser linfocitos T que expresan, además del CD3 con receptor de linfocito T-αβ, receptores propios de células NK como CD161. Reconocen glucolípidos presentados en el contexto de CD1d. Su función tolerogénica no se conoce lo suficiente, si bien parece estar mediada por interacciones con otras células reguladoras y la secreción de citocinas moduladoras de la respuesta, como IL-4, IL-10 y TGF-â43.
- •
Los linfocitos B reguladores. La función reguladora de la respuesta inmune por parte de los linfocitos B se describió hace más de 40 años, y se atribuyó entonces a una supuesta capacidad de producir anticuerpos inhibitorios44. Los linfocitos Br (B reguladores) han sido identificados en relación con la pérdida de la tolerancia en modelos experimentales de autoinmunidad, infecciones y cáncer, y son fácilmente identificables debido a que son CD1d+hi y CD5+45. Su actividad reguladora parece estar mediada por un doble mecanismo: la producción de citocinas inmunomoduladoras como IL-10 y TGF-β, y la capacidad de interactuar directamente con linfocitos T reactivos, a los que son capaces de inhibir46.
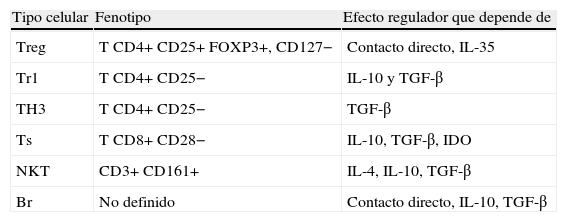
Células T reguladoras
| Tipo celular | Fenotipo | Efecto regulador que depende de |
| Treg | T CD4+ CD25+ FOXP3+, CD127− | Contacto directo, IL-35 |
| Tr1 | T CD4+ CD25− | IL-10 y TGF-β |
| TH3 | T CD4+ CD25− | TGF-β |
| Ts | T CD8+ CD28− | IL-10, TGF-β, IDO |
| NKT | CD3+ CD161+ | IL-4, IL-10, TGF-β |
| Br | No definido | Contacto directo, IL-10, TGF-β |
Br: linfocitos B reguladores; IDO: indolamina-2,3-dioxigenasa; IL: interleucina; NKT: linfocitos T citolíticos naturales; TGF-β: factor del crecimiento transformante β; TH: linfocito T helper; Treg: linfocitos T reguladores; Tr1: linfocitos T reguladores inducibles; Ts: linfocitos T supresores.
El paradigma de las células reguladoras como guardianes de la tolerancia a lo propio como consecuencia de su actividad en la supresión de la respuestas inmunes implica que su hiperfunción debe acarrear una menor respuesta a la infección. Este paradigma ha sido recientemente torpedeado por un magnífico trabajo del grupo de Rudensky, que ha demostrado el papel fundamental de las células reguladoras como colaboradoras necesarias en los primeros momentos de la respuesta inmune. Estos autores, mediante el estudio de un modelo murino de infección letal por virus del herpes en mucosas, observaron que la carga vírica era mayor, y la progresión de la infección era mucho más rápida en los ratones que carecían de Treg47.
Este trabajo demuestra que las Treg son esenciales en las respuestas tempranas a la infección vírica local y facilitan la entrada de células inmunes (NK, TH17) en el tejido infectado. Esta actividad es indudablemente de tipo colaborador, hecho que debe hacer reflexionar acerca de su papel y considerarlas, al igual que a las colaboradoras, como coordinadoras de la respuesta inmune (el título del trabajo ya lo sugiere).
Una célula coordinadora tendría, a la vez, funciones colaboradoras y supresoras de la respuesta, promoviendo unas acciones e inhibiendo otras. Este fenómeno se demuestra claramente con las TH1, TH2 y TH17. Además de colaboradoras, tienen funciones supresoras de las otras respuestas, ya que son mutuamente antagónicas.
A este grupo de células que actúan tanto de forma colaboradora como reguladora se les unirían también las Treg, ya que además de su conocida función supresora también coordinarían los primeros pasos de la infección vírica de modo directo como también indirectamente induciendo, mediante la secreción de TGF-β, la diferenciación de las TH17.


